
小麦是一种主要的粮食作物,为人类提供大约四分之一的食物热量。小麦产量的持续增长对应对粮食消费增长的挑战至关重要。小麦植物株型包括株高、分蘖数、分蘖角,叶片大小和叶角,这些性状都显著影响小麦产量。旗叶形态包括长、宽、面积、叶基角、叶开角和披垂度等性状。虽然已经有许多关于旗叶的QTL已被检测到,但仅少数稳定表达的主效位点被报道。
小麦品系20828具有小穗数多()、株高适中和高抗条锈()等优异性状,近十年被广泛用于育种亲本。川农16是国审小麦品种,具有株型优异、分蘖多特性()。前期我们基于20828×川农16重组自交系群体,对二者控制旗叶相关性状的位点进行了挖掘,揭示了川农16优异株型的遗传基础。品系SY95-71,易感条锈病,但具有株叶型好、分蘖多()、穗颈适中()、根系发达()等优良特性,近十年也一直被用作育种亲本。为了进一步解析SY95-71株叶型的遗传基础,我们基于20828×SY96-71(2SY,F7)重组自交系群体、小麦55KSNP芯片构建的遗传图谱、多环境和多群体对旗叶形态相关性状位点进行了挖掘、鉴定和验证。

图1亲本和部分后代株系的旗叶表型
结果
1.旗叶表型分析
如图1所示,亲本20828的叶基角(FLANG)、叶开角(FLOA)和披垂度(FLBA)显著高于SY95-71,并且重组自交系群体中旗叶相关性状的表型值都呈连续分布趋势(图2)。旗叶长(FLL)、宽(FLW)、面积(FLA)、基角、开角和披垂度的遗传力(h2)分别为0.60、0.66、0.62、0.57、0.69和0.68。
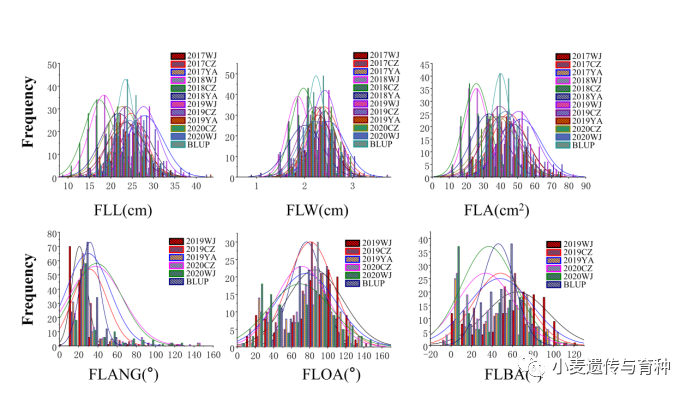
图2重组自交系群体旗叶表型分布
2.旗叶与产量性状的相关性分析
旗叶性状之间的相关性分析结果表明:旗叶长与旗叶宽、面积、开角以及披垂度呈显著正相关();旗叶面积分别与旗叶宽、开角以及披垂度呈显著正相关()(图3)。
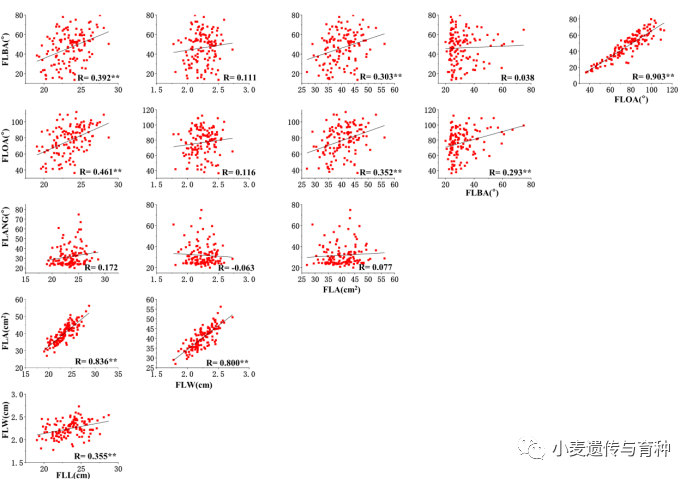
图3旗叶性状之间的相关性分析
旗叶性状与产量性状之间的相关性分析表明:旗叶宽与小穗数(SNS)、粒宽(GW)以及千粒重(TGW)呈显著正相关();旗叶面积与千粒重呈显著正相关();旗叶开角与千粒重呈显著正相关();披垂度与粒长(GL)及千粒重呈显著正相关();旗叶基角与粒宽呈显著负相关();旗叶开角角与小穗数呈显著负相关();披垂度与小穗数呈显著负相关()。
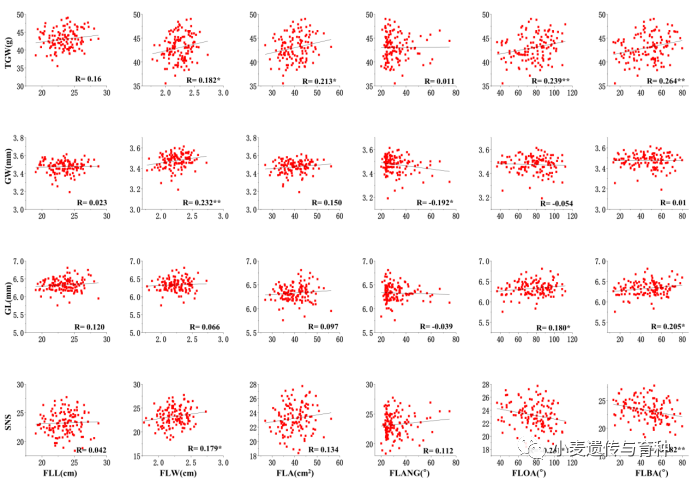
图4旗叶性状与产量性状之间的相关性分析
3.旗叶QTL检测
本研究采用完备区间作图法,在11个环境中共检测到86个和旗叶形态相关的QTL,这些QTL中有13个旗叶长QTL,10旗叶宽QTL,27个旗叶面积QTL,7个旗叶基角QTL,14个旗叶开角QTL和15个披垂度QTL。有8个QTL是主效的,能够在超过4个环境中表达,此外,有78个只在3个环境及以下被检测到,不能稳定表达。
本研究在11个环境中一共定位了13个控制旗叶长的QTL。13个QTL分别位于1B、1D、2B、2D、3D、4D、5B、7A和7B染色体上,LOD值在2.54~28.88之间,可解释的表型变异在2.88%和54.38%之间。2个QTL能在超过4个环境中被检测到,且贡献率大于10%,起主效效应,将它们分别命名为和。位于2B染色体上AX-109331624~AX-109893758区间内,对旗叶长的加性效应分别来自SY95-71(图5);位于5B染色体上AX-109443553~AX-108867784区间内,对旗叶长的加性效应分别来自20828。
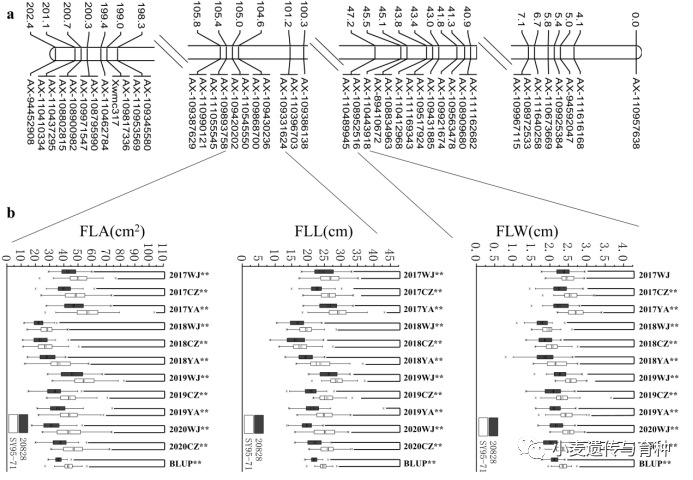
图5、和的遗传图谱
本研究获取的10个旗叶宽QTL分别分布在1A、2B、2D、3A、3B、4B、4D、6B和7A染色体上,它们的LOD值介于2.51和25.40之间,表型贡献率在4.08%和48.89%之间。有两个QTL是主效的,分别命名为和,位于2B染色体上AX-110443918~AX-108952516区间内,在9个环境中被检测到,可解释的表型变异是15.05%~48.89%,加性效应来自亲本SY95-71(图5);位于2D染色体上AX-109983215~AX-109402447区间内,在4个环境中被检测到,可解释的表型变异是8.89%~19.06%,加性效应来自亲本20828。
27个旗叶面积QTL中,有一个稳定表达的QTL()位于2B染色体上AX-109868700~AX-109893758区间内,其加性效应来自SY95-71,解释了5.73-31.74%的表型变异。其余的QTL分布在1D、2B、2D、3D、4D、5A、5B、6B、6D、7A和7B染色体上,它们的LOD值介于2.57和14.64之间,解释了2.64-40.49%的表型变异。
本研究一共检测到7个旗叶基角QTL,其减小旗叶基角的加性效应都来自SY95-71,其中有一个稳定表达的QTL,命名为,位于4B染色体上AX-111233094~AX-111168324区间内,解释9.51-35.87%的表型变异;剩余的QTL分布在1D、4D、5A、5B和7B染色体上,它们的LOD值介于2.54和4.38之间,解释了2.24%-8.17%的表型变异。
在本研究检测到14个旗叶开角QTL中,有一个稳定表达的QTL,命名为,位于染色体上AX-109400957~AX-111716941区间内,其减小旗叶开角的加性效应来自SY95-71,解释14.60-26.24%表型变异。剩余的13个QTL分布在1A、2B、2D、3A、3D、4A、5D、7A和7B染色体上,它们的LOD值介于2.64和11.15之间,解释了4.76-23.22%表型变异。
在检测到的15个控制披垂度QTL中,有一个稳定表达的QTL,命名为,也位于6D染色体上AX-109400957~AX-111716941区间内,其减小旗叶开角的加性效应也来自SY95-71,解释11.95-27.46%表型变异。剩余的14个QTL分布在1A、1B、2A、2B、3A、4A、5D、6B、7A、7B和7D染色体上,它们的LOD值介于2.54和13.55之间,它们解释了3.77-23.57%的表型变异。
在这8个主效QTL中,和共定位于AX-109331624~AX-109893758之间(图5),和共定位于AX-109400957~AX-111716941之间(图6)。
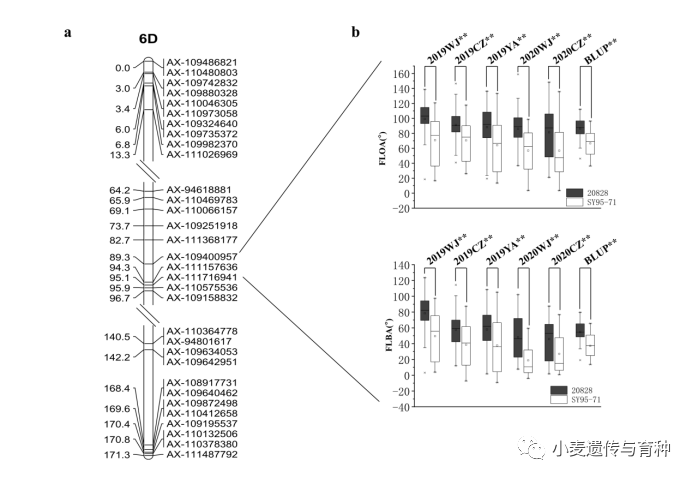
图6和的遗传图谱
4.2SY群体中两个主效QTL对旗叶长和宽的影响
和的加性效应分别来自SY95-71和20828,进一步分析了它们对旗叶长的影响(图7a)。与不携带有任何位点的株系相比,仅携带或仅携带位点的株系,可以分别显著性增加12.21%或7.70%的旗叶长;而同时携带这两个位点的株系可以显著性增加20.91%的旗叶长。此外,同时携带这两个位点的株系旗叶长比只携带或位点的株系旗叶长增加了7.76%或12.27%。
和的加性效应分别来自SY95-71和20828,它们对旗叶宽的影响如图7b所示。与不携带有任何位点的株系相比,仅携带或仅携带位点的株系,可以分别显著性增加8.27%或5.87%的旗叶宽;而同时携带这两个位点的株系可以显著性增加13.96%的旗叶宽;此外,同时携带这两个位点的株系旗叶宽比只携带或位点的株系旗叶宽增加了5.25%或7.64%。
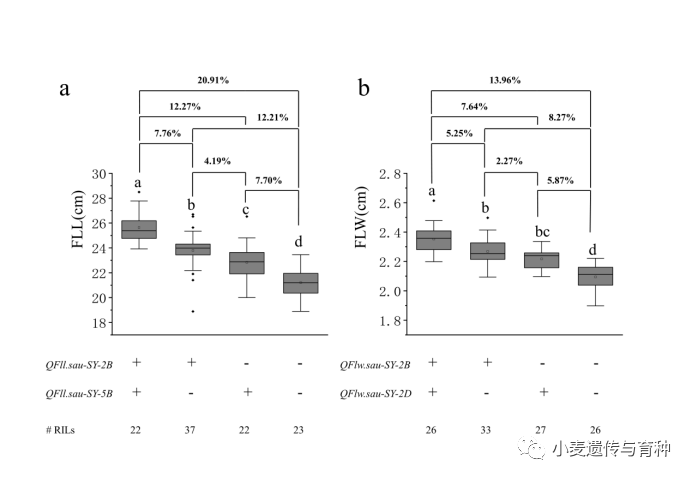
图7两个主效QTL对旗叶长和宽的影响
5.主效QTL对产量性状的影响
在2SY群体中,进一步分析了主效QTL对产量性状的影响,携带、和的株系与不携带这些位点的株系之间的小穗数、粒长、粒宽和千粒重差异不显著(图8a)。同时携带和的株系,与不携带任何位点的株系相比显著增加1.25%的粒宽和4.69%的千粒重,与仅携带的株系相比显著增加3.67%的千粒重(图8b)。携带的株系与不携带任何位点的株系相比显著增加3.19%的小穗数(图8c)。携带/的株系与不携带任何位点的株系相比显著增加1.61%的粒长、1.28%的粒宽和3.90%的千粒重(图8d)。
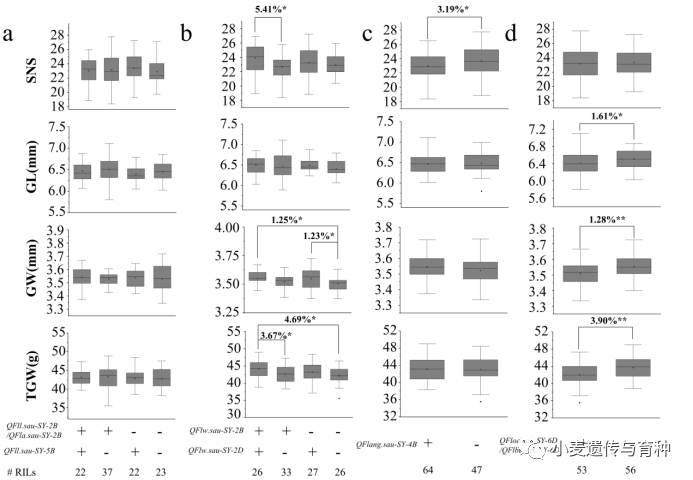
图8主效QTL对产量性状的影响
6.在不同遗传背景中验证
在检测到的8个主效QTL中,和可能和我们之前研究中所检测到的和是相同的位点,这两个QTL也在不同遗传背景群体中能够稳定表达()。
我们利用新开发的KASP标记KASP-AX-109331624在6个不同遗传背景的群体(SSY、KS、4S、5S、8S和10S)中去验证和的效应。根据标记的在群体中分型结果,将群体分为两组(排除杂合型),再对两组进行独立样本t检验。正如预期的那样,t检验发现他们之间存在显著性差异(),携带SY95-71的等位基因株系的旗叶长和面积显著高于不携带该等位基因株系,旗叶长差异分别是9.05%、12.48%、9.60%、8.46%、9.31%和7.44%(图9a);旗叶面积差异分别是9.03%、20.35%、20.53%、24.30%和18.42%(图9b)。
利用与连锁的KASP标记(KASP-AX-110443918和KASP-AX-108952516)在上述的6个群体中验证。结果表明携带SY95-71的等位基因的株系显著高于不携带该等位基因株系的旗叶宽,差异分别是7.34%、7.45%、12.55%、6.37%、12.92%和7.00%(图9c)。
因为在以往的研究中表明直立叶对于小麦产量提升更为显著,所以本研究在不同的遗传背景中验证其减小旗叶角度的效应。
对于,新开发KASP标记KASP-AX-109110130只在SSY、4S和8S群体中检测到多态性。结果表明携带SY95-71的等位基因株系的旗叶基角显著小于不携带该等位基因株系,差异分别是38.86%、57.73%和11.90%(图9d)。
和的验证结果表明:在SSY、KS和5S群体中,携带SY95-71的等位基因株系的旗叶开角显著小于不携带该等位基因的株系,差异分别是23.38%、22.53%和38.87%(图9e);在SSY、KS、5S、8S和10S群体中,携带SY95-71的等位基因株系披垂度显著小于不携带该等位基因的株系,差异分别是38.64%、33.42%、58.94%、33.99%和39.62%(图9f)。
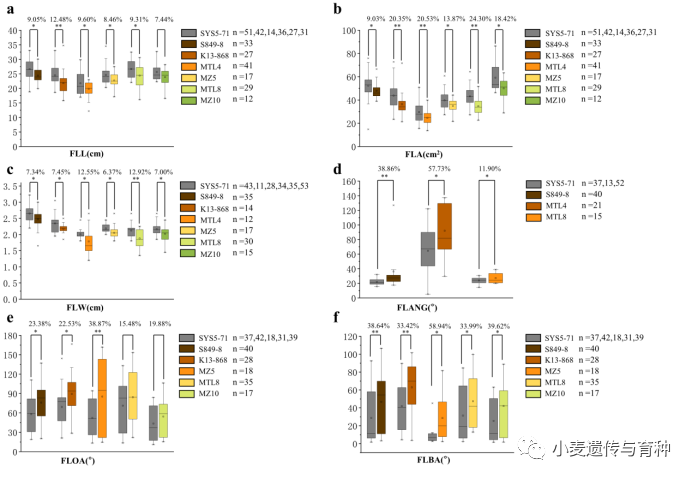
图9在不同遗传背景群体中的验证
7.新的QTL
本研究中检测到的8个主效QTL中,、和都可能是和我们之前的研究中所检测的QTL是相同的位点()。位于2B染色体634.4Mbp和663.3Mbp之间,在该区间附近没有与旗叶长相关QTL被报道,所以可能是一个新的位点。其他的位点的物理区间附近均有相关的QTL被报道的,表明这些QTL可能不是新的位点。
讨论
1.在不同的遗传背景下检测不同QTL
在利用20828×川农16构建的重组自交系检测旗叶长、宽、面积、基角、开角和披垂度相关QTL,检测到4个主效位点:、共定位位点//、和共定位位点/,这些位点都由20828所贡献。本研究中(20828×SY95-71),我们在的区间附近也检测到;共定位位点//的区间附近检测到;的区间附近检测到。但是没有在2D染色体检测到与旗叶角度相关的QTL。造成这一结果的原因可是由于20828和SY95-71之间在/的区间内没有等位基因之间的变异。
2.候选基因的预测
利用和的侧翼标记的序列在中国春和野生二粒参考基因组上进行Blast,获得该位点的在参考基因组上的物理区间,然后再从数据库获得两个区间的参考基因,最后把两组基因进行分析获得同源基因。通过查找文献寻找与叶发育相关的基因,如、、等,这些基因在植物中调控叶的生长发育,很可能就是我们的候选基因。同时,我们利用中国春和节节麦的参考基因组对/区间内的基因进行分析,找到一些可能的候选基因,如、、等。
通过数据库获得候选基因的表达模式(图10),选取了在叶中表达量较高的两个基因(TraesCS2B02G457600和TraesCS2B01G458800)进行克隆测序。结果表明,他们的序列828和SY95-71之间存在序列差异,这些差异为进一步确定目的基因奠定基础。
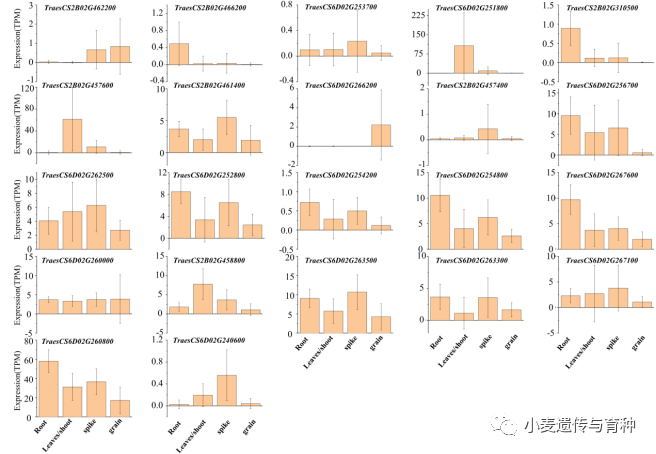
图10候选基因的表达
3.QTL对目标性状的改善
以往的研究认为,优异基因的叠加是改善目标性状的有效策略。利用重组和标记辅助选择技术,将多个优良QTL重组到一个新品种上可以进一步改善目标性状。在本研究中,我们发现和对旗叶长的影响强烈,和对旗叶宽的影响显著,聚合这些加性位点对进一步调控旗叶长和宽,从而改善株型具有一定意义。
该研究相关结果于2020年10月7日发表在TheoreticalandAppliedGenetics杂志上(QTLmappingandvalidationofbreadwheatflagleafmorphologyacrossmultipleenvironmentsindifferentgeneticbackgrounds(2020))。
该研究得到国家自然科学基金、四川省重点研发项目、四川省科技厅应用基础研究项目、四川省留学生科技活动重点项目的资助。
主要参考文献:
LiuJ,TangH,QuX,LiuH,LiC,TuY,LiS,HabibA,MuY,DaiS,DengM,JiangQ,LiuY,ChenG,WangJ,ChenG,LiW,JiangY,WeiY,LanX,ZhengY,MaJ(2020)Anovel,major,andvalidatedQTLf:173-185
MaJ,TuY,ZhuJ,LuoW,LiuH,LiC,LiS,LiuJ,DingP,HabibA,MuY,TangH,LiuY,JiangQ,ChenG,WangJ,LiW,PuZ,ZhengY,WeiY,KangH,ChenG,LanX(2020)Flagleafsizeandpostureofbreadwheat:geneticdissection,:297-315
MaJ,DingP,LiuJ,LiT,ZouY,HabibA,MuY,TangH,JiangQ,LiuY,ChenG,WangJ,DengM,QiP,LiW,PuZ,ZhengY,WeiY,XL(2019a)Identificationandvalidationofa:3155-3167
MaJ,QinN,CaiB,ChenG,DingP,ZhangH,YangC,HuangL,MuY,TangH,LiuY,WangJ,QiP,JiangQ,ZhengY,LiuC,WeiY(2019b)IdentificationandvalidationofanovelmajorQT:1363-1373
LiC,TangH,LuoW,ZhangX,MuY,DengM,LiuY,JiangQ,ChenG,WangJ,QiP,PuZ,JiangY,WeiY,ZhengY,LanX,MaJ(2020)Anovel,validated,andplantheight-indepentQTLfo:
LiuJ,LuoW,QinN,DingP,ZhangH,YangC,MuY,TangH,LiuY,LiW,JiangQ,ChenG,WeiY,ZhengY,Liuc,LanX,MaJ(2018)A55KSNParray-basedgeneticmapandi:2439-2450
ZhengX,WenX,QiaoL,ZhaoJ,ZhangX,LiX,ZhangS,YangZ,ChangZ,ChenJ(2019)ionanalysesinwheat(TriticumaestivumL.).Planta250:129-143

扫描二维码
小麦族多组学网站:
投稿、合作等邮箱:@163.com







